Lajinmuodostus: miten uudet lajit syntyvät — anageneesi, kladogeneesi
Tutustu lajinmuodostukseen: anageneesi vs. kladogeneesi, hybridisaation rooli ja nykytutkimuksen löydöt evoluution mekanismeista.
Lajistumisessa on kyse siitä, miten lajit muodostuvat. Se on tärkeä osa evoluutiobiologiaa. Lajistuminen selittää, miten luonnon monimuotoisuus on syntynyt ja millä mekanismeilla populaatioista eriytyy uusia, toisistaan erottuvia evolutiivisia linjoja.
Kuvagalleria
10 Kuvat









Anageneesi ja kladogeneesi
Darwin ajatteli, että useimmat lajit ovat peräisin suoraan jo olemassa olevista lajeista. Tätä kutsutaan nimellä anagenesis: lajit muuttumalla tai "fyletic evolution". Anageneesissä koko populaatio muuttuu ajan myötä siten, että alkuperäinen muoto muuttuu uuteen ilman selkeää jakautumista kahdeksi erilliseksi lajiksi.
Sitä vastoin kladogeneesi tarkoittaa tilannetta, jossa yksi populaatio jakautuu kahdeksi tai useammaksi erilliseksi linjaksi, joista muodostuu uusia lajeja. Kladogeneesi on monille tutumpaa mallia: yksi kantamuoto antaa syntymän useille perillislajeille.
Miten lajittuminen tapahtuu — mekanismit
Lajinmuodostuksen taustalla vaikuttavat useat biologiset prosessit. Tärkeitä tekijöitä ovat luonnonvalinta, sattumanvarainen geenivirta (genetic drift), mutaatiot, geneettinen eristys ja populaation rakenteen muutokset (esim. perustajavaikutus).
Yleisten mekanismien ryhmittely:
- Maantieteellinen eristäytyminen (allopatrinen lajinmuodostus): populaatio jakautuu fyysisesti erillisiksi osiksi (esim. vuoristo, jäätikkö, saaret). Eristäytyminen estää tai vähentää geenivirtaa, jolloin paikallinen sopeutuminen ja sattuma johtavat eroihin.
- Peripatrinen lajinmuodostus: pieni populaatio eristyy reuna-alueelle tai uudelle ekologiselle tilalle (esim. saarelle). Pieni koko ja perustajavaikutus voivat nopeuttaa geneettisten erojen kertymistä.
- Parapatrinen lajinmuodostus: populaation osuudet ovat vierekkäin ja lisääntymistä tapahtuu rajassa, mutta erilaistuminen voi syntyä erilaisista ekologisista valintapaineista ja vähentyneestä geenivirrosta.
- Sympatrinen lajinmuodostus: uudet lajit syntyvät samalta alueelta ilman selkeää fyysistä estettä. Tyypillisiä mekanismeja ovat vahva ekologinen erikoistuminen, seksuaalivalinta tai kromosomipoikkeamat kuten polyploidia (yleistä kasveilla).
Lisääntymiseristys — pre- ja postzygoottiset esteet
Lajinmuodostus edellyttää yleensä jonkinasteista lisääntymiseristystä. Se voidaan jakaa:
- Prezygoottiset esteet — estävät risteytymisen ennen hedelmöitystä: maantieteellinen erillisyys, eriajoitus (temporal), käyttäytymiserot (esim. soidinlaulu), mekaaninen sopimattomuus ja gametinen eristys.
- Postzygoottiset esteet — vaikuttavat risteytyksen jälkeen: hybridien heikentynyt eloonjäänti, kasvava steriilisyys tai heikentynyt lisääntyminen (esim. muulit ovat steriilejä).
Hybridisaatio ja geenivirta
Viimeisten vuosikymmenten tutkimukset, erityisesti elävien olentojen DNA-sekvenssin analysointi, ovat osoittaneet, että sukulaislajien välillä esiintyy usein jonkin verran hybridisaatiota. Tämä tarkoittaa, että geenit voivat siirtyä lajien välillä ja vaikuttaa niiden evoluutioon. Tästä seuraa kaksi tärkeää tulkintaa:
- Lisääntymiseristys ei välttämättä ole täydellinen lajin rajan määrittäjä — lajien välillä voi olla geenivirtaa pitkäänkin.
- Hybridisaatio voi tuoda populaatioihin uusia perintötekijöitä, jotka nopeuttavat sopeutumista uusiin olosuhteisiin tai synnyttävät täysin uusia geneettisiä yhdistelmiä, mikä joskus edistää uuden lajin syntyä.
Erityistapauksia: kasvien polyploidia ja nopea speciaatio
Kasveilla polyploidia (kromosomiston kaksoistuminen tai moninkertaistuminen) on yleinen ja nopea tapa muodostaa lisääntymisesteisiä populaatioita. Yksi polyploidinen yksilö voi nopealla aikaskaalailla muodostaa uuden, toisesta äidinlajista erottuvan populaation.
Esimerkkejä ja havainnollistuksia
Monia esimerkkejä lajinmuodostuksesta on kuvattu tutkimuskirjallisuudessa:
- Darwinin tikat (esimerkki sopeutumisesta saariston eri ruokavalioihin ja nokan muotoihin) — antaa kuvan, miten eri ekologinen valinta voi ajaa lajiutumista.
- Ring-lajit (esim. jotkin lokkilajiryhmät tai salamanderi Ensatina): populaatio muodostaa rengasmaisesti jatkuneen jakauman, jonka vastakkaiset päät mukautuvat eri tavoilla ja eivät enää lisäänty kovin hyvin keskenään.
- Rhagoletis-puuvaarapistiäinen (omena vs. pihlaja): esimerkki sympatrisesta tai parapatrisesta lajinmuodostuksesta, jossa isäntäkasvin vaihtuminen johti eriytymiseen.
- Afrikan endeemiset cichlidit: uskomaton lajirunsaus järvissä, jossa ekologinen erikoistuminen ja seksuaalivalinta ovat johtaneet moniin lajeihin lyhyessä ajassa.
Lajinmuodostuksen aikaskaala ja nykymalli
Spektri on laaja: jotkin lajit muuttuvat hitaasti anageneesin kautta, toiset eriytyvät nopeasti kladogeneesin kautta. Moderni tutkimus korostaa speciation-kontinuumia: populaatioiden väliset erot kehittyvät asteittain pre- ja postzygoottisten esteiden vahvistuessa. Lisäksi nykyiset genomitutkimukset osoittavat usein monimutkaisia verkostoja, joissa lajit eivät ole täysin haarautuneita puumaisesti vaan risteävät ajoittain toistensa kanssa.
Päätelmä
Lajinmuodostus ei ole yksi yksinkertainen prosessi vaan joukko erilaisia mekanismeja ja reittejä, joilla populaatiot eriytyvät geneettisesti ja lisääntymisesti. Sekä anagenesis että kladogeneesi ovat tärkeitä käsitteitä, joiden merkitys riippuu tilanteesta. Fyysinen erillisyys on usein keskeinen tekijä, mutta genomi- ja kenttätutkimukset ovat osoittaneet, että myös hybridisaatio, geneettinen vaihtelu ja nopea kromosomimuutos voivat olla ratkaisevia uusien lajien syntymisessä.
Aihetta koskeva tutkimus jatkuu aktiivisesti, ja modernit DNA-analyysit sekä kenttihavainnot auttavat ymmärtämään lajinmuodostuksen monimuotoisuutta ja dynamiikkaa entistä paremmin.
Eristysmekanismit
Eristämismekanismit ovat asioita, jotka estävät lajin ryhmien välisen lisääntymisen onnistumisen. Populaatioiden lisääntymiseristys on vakiintunut. Tämä on erityisen tärkeää biologisen lajikäsitteen kannalta, sillä lajit määritellään lisääntymiseristyksen perusteella.
Eristysmekanismit voidaan jakaa kahteen ryhmään, ennen ja jälkeen hedelmöityksen.
Ennen lannoitusta
Tekijät, jotka estävät yksilöitä pariutumasta.
- Maantieteellinen eristyneisyys: Lajit esiintyvät eri alueilla, ja ne ovat usein esteiden erottamia.
- Ajallinen eristäminen: Yksilöt eivät pariudu, koska ne ovat aktiivisia eri aikoina. Tämä voi olla eri vuorokaudenaikoja tai eri vuodenaikoja. Lajien pariutumisajat eivät välttämättä sovi yhteen. Yksilöt eivät tapaa toisiaan paritteluaikoina tai lainkaan.
- Ekologinen eristäminen: Yksilöt parittelevat vain haluamassaan elinympäristössä. Ne eivät kohtaa muiden lajien yksilöitä, joilla on erilaiset ekologiset mieltymykset.
- Käyttäytymisen eristäminen: Eri lajeja edustavat yksilöt voivat tavata, mutta toinen ei tunnista mitään seksuaalisia vihjeitä. Yksilö valitsee useimmiten oman lajinsa jäsenen.
- Mekaaninen eristys: Kopulaatiota voidaan yrittää, mutta siittiöiden siirtoa ei tapahdu. Yksilöt voivat olla yhteensopimattomia koon tai morfologian vuoksi.
- Geneettinen yhteensopimattomuus: Spermansiirto tapahtuu, mutta munasolu ei hedelmöity.
Lannoituksen jälkeen
Tekijät, jotka estävät parittelun onnistumisen, kuten geneettinen yhteensopimattomuus, hybridin elinkyvyttömyys tai steriiliys.
- Zygoottinen kuolleisuus: Muna hedelmöittyy, mutta sikiö ei kehity.
- Hybridin elinkyvyttömyys: Hybridi alkio muodostuu, mutta ei ole elinkelpoinen.
- Hybridisteriliteetti: Hybridi on elinkelpoinen, mutta syntyvä aikuinen on steriili.
- Hybridi jaottelu: Ensimmäisen sukupolven (F1) hybridit ovat elinkelpoisia ja hedelmällisiä, mutta myöhemmät hybridisukupolvet (F2 ja takaisinristeytykset) ovat elinkelvottomia tai steriilejä.

Maantieteellinen eristyneisyys
Tämän uskotaan olevan yleisin syy lajinmuodostukseen. Ensimmäisenä sen keksi saksalainen tutkimusmatkailija ja luonnonhistorioitsija Moritz Wagner.
Wagnerin uran alkuvaiheessa hän toimi maantieteilijänä ja julkaisi useita maantieteellisiä kirjoja Pohjois-Afrikasta, Lähi-idästä ja trooppisesta Amerikasta. Hän oli myös innokas luonnontieteilijä ja keräilijä, ja biologit tuntevat hänet parhaiten juuri tästä työstään. Evoluutiotieteilijä ja biologian historioitsija Ernst Mayr on kertonut Wagnerin merkityksestä.p562–565
Kolmen Algeriassa viettämänsä vuoden aikana Wagner tutki muun muassa lentokyvyttömiä kuoriaisia Pimelia ja Melasoma. Kumpikin suku jakautuu useisiin lajeihin, joista kukin on rajoittunut Atlasvuorilta Välimerelle laskevien jokien väliselle pohjoisrannikolle. Heti kun joki ylitetään, esiintyy eri laji, joka on kuitenkin läheistä sukua.
"... alkava laji [syntyy] vain silloin, kun muutamat yksilöt ylittävät levinneisyysalueensa rajat... uuden rodun muodostuminen ei koskaan onnistu... ilman, että siirtolaiset erottautuvat pitkään muista lajinsa jäsenistä".
Tämä oli varhainen kuvaus eräänlaisesta maantieteellisestä lajistumisprosessista. Mayr otti sen uudelleen käyttöön vuonna 1942, ja maantieteellisen lajiston merkityksestä tuli yksi evoluutiosynteesin ydinajatuksista.
Toinen termi maantieteelliselle lajistumiselle on allopatrinen lajistuminen. Allopatria tarkoittaa 'eri maata'.
Tyhjät saaret
Tuliperäiset saaret muodostuvat ilman elämää, ja kaiken elämän on tultava tuulen tai veden mukana. Havaijin saarilta ja Galapagossaarilta tiedämme, että kaikki elämänmuodot muuttuvat, kun ne saapuvat saarille mantereelta.
Havaijinsaarilla on noin 17 000 kilometrin (6 500 neliömetrin) alueella (2 ) maailman monipuolisin kokoelma drosofilidikärpäsiä, jotka elävät sademetsistä vuoristoniittyihin. Havaijilaisia drosofilidilajeja tunnetaan noin 800.
Tutkimukset osoittavat, että lajeja siirtyy selvästi vanhemmilta saarilta uusille saarille. On myös tapauksia, joissa lajit siirtyvät takaisin vanhemmille saarille ja jättävät saaria väliin, mutta nämä ovat paljon harvinaisempia.
Kalium/argon-radioaktiivisen ajanmäärityksen mukaan nykyiset saaret ovat peräisin 0,4 miljoonaa vuotta sitten (mya) (Mauna Kea) ja 10 miljoonaa vuotta sitten (Necker). Havaijin saariston vanhin vielä merenpinnan yläpuolella oleva osa on Kure-atoll, joka voidaan ajoittaa 30 mya:n ikäiseksi.
Itse saaristo, joka on syntynyt Tyynenmeren laattojen liikkuessa kuuman pisteen yli, on ollut olemassa paljon kauemmin, ainakin liitukaudella. Havaijin saaret ja entiset saaret, jotka ovat nyt meren alla, muodostavat Havaijin ja Keisarin saarien ketjun, ja monet vedenalaisista vuorista ovat guyotteja.
Kaikki Havaijin kotoperäiset drosofilidilajit ovat ilmeisesti peräisin yhdestä esi-isälajista, joka asutteli saaret noin 20 miljoonaa vuotta sitten. Tätä seuranneen sopeutumissäteilyn taustalla oli kilpailun puute ja monenlaiset vapaat markkinaraot. Vaikka yksittäinen tiineenä oleva naaras olisi voinut asuttaa saaren, on todennäköisempää, että kyseessä on ollut saman lajin ryhmä.
Havaijin saaristossa on muitakin eläimiä ja kasveja, jotka ovat kokeneet samanlaisia, joskin vähemmän näyttäviä sopeutumissäteitä.
Rengaslajit
Biologiassa rengaslajilla tarkoitetaan vierekkäisten populaatioiden yhdistettyä sarjaa, joista kukin voi risteytyä naapuripopulaatioiden kanssa. Ketjun kaksi päätä ovat päällekkäin.
Sarjan kaksi loppupopulaatiota ovat liian kaukana toisistaan, jotta ne voisivat risteytyä keskenään. Tällaiset ei-lajittuvat, mutta geneettisesti toisiinsa yhteydessä olevat "loppupopulaatiot" voivat elää samalla alueella rinnakkain, jolloin "rengas" sulkeutuu.
Rengaslajit ovat tärkeä todiste evoluutiosta: ne havainnollistavat, mitä ajan kuluessa tapahtuu, kun populaatiot eroavat geneettisesti toisistaan. Richard Dawkins huomautti, että rengaslajit "näyttävät meille vain alueellisessa ulottuvuudessa jotakin, minkä on aina tapahduttava aikaulottuvuudessa".
On kuitenkin vaikea löytää yksinkertaista ja suoraviivaista esimerkkiä.
Larus-lokit
Tämä oli klassinen esimerkki rengaslajeista. Näiden lokkien levinneisyysalue muodostaa lähes renkaan pohjoisnavan ympärille (jonka yli lokit eivät yleensä lennä). Lokkilokit ja silakkalokit ovat niin erilaisia, että ne eivät yleensä risteydy keskenään; sanottiin, että lokiryhmä muodostaa jatkumon paitsi siellä, missä nämä kaksi sukulinjaa kohtaavat Euroopassa. Yleisesti ollaan kuitenkin yhtä mieltä siitä, että tämä ei pidä aivan paikkaansa, vaikka yksityiskohdat ovatkin erittäin monimutkaisia.
Ensatina-salamanterit
Ensatina-salamanteri on rengaslaji Kalifornian Keskilaaksoa ympäröivillä vuorilla. Kompleksi muodostaa hevosenkengän muodon vuorten ympärille. Vaikka hevosenkengän ympärillä olevat 19 populaatiota voivat risteytyä keskenään, hevosenkengän länsipäässä oleva Ensatina eschscholtzii -alalaji ei voi risteytyä itäpäässä olevan Ensatina klauberi -alalalajin kanssa. Se on esimerkki "lähes kaikista lajinmuodostumisprosessin vaiheista" (Dobzhansky). Richard Highton väitti, että Ensatina on usean lajin tapaus eikä yhden lajin jatkumo.
Vihertävä laulurastas
Vihervarpulla (Phylloscopus trochiloides) on useita alalajeja, joista P. t . viridianus on Euroopassa tunnetuin. Se on rengaslaji, jonka populaatiot eroavat toisistaan Tiibetin ylängön itä- ja länsipuolella ja kohtaavat myöhemmin pohjoispuolella. Niiden väliset suhteet ovat melko sekavat.




Sympatrinen lajisto
Sympatrinen lajisto tarkoittaa kahden tai useamman jälkeläislajin muodostumista yhdestä esi-isälajista, jotka kaikki asuvat samalla maantieteellisellä alueella. Tämän uskotaan nykyään olevan hyvin yleistä.
Sympatrisessa lajinmuodostuksessa lajit eroavat toisistaan asuessaan samassa paikassa. Usein mainittuja esimerkkejä sympatrisesta lajistumisesta ovat hyönteiset, jotka tulevat riippuvaisiksi eri isäntäkasveista samalla alueella.
Sympatrisen lajinmuodostuksen olemassaolosta lajinmuodostuksen mekanismina kiisteltiin kiivaasti. Väitettiin, että sympatrisen lajiston todisteet ovat itse asiassa esimerkkejä mikrogeografisesta lajistumisesta. Yleensä tämä ei ole nykyään suosituin selitys. Pikemminkin se on seurausta läheisesti sukua olevien lajien välisestä risteytymisestä, jonka jälkeen luonnonvalinta vaikuttaa tällaisten risteytysten jälkeläisiin. Yksi laajalti hyväksytty esimerkki sympatrisesta lajistosta on Itä-Afrikassa sijaitsevan Nabugabo-järven sirkkalajien lajisto, jonka uskotaan johtuvan sukupuolisesta valinnasta.
Lajittuminen polyploidisaation kautta
Polyploidia on aiheuttanut monia nopeita lajinmuodostustapahtumia, koska esimerkiksi tetraploidi x diploidi -paritusten jälkeläiset ovat usein triploideja steriilejä jälkeläisiä.
Kaikki polyploidit eivät kuitenkaan ole lisääntymiseristettyjä emokasveistaan, ja geenivirtaa voi edelleen esiintyä esimerkiksi triploidien hybridien ja diploidien parittelussa, joka tuottaa tetraploideja.
Monet nykyiset kasvi- ja useimmat eläinlajit ovat ilmeisesti käyneet läpi polyploidisaation evoluutiohistoriansa aikana. Menestyneiden polyploidisten lajien lisääntyminen on joskus suvutonta, parthenogeneesin avulla. Tuntemattomista syistä monet suvuttomat eliöt ovat polyploideja.
Hawthorn kärpässieni
Yksi esimerkki evoluution toiminnasta on orapihlajakärpänen Rhagoletis pomonella, joka näyttää olevan sympatrisen lajinmuodostuksen kohteena.
Orapihlajakärpäsen eri populaatiot syövät eri hedelmiä. Erillinen populaatio syntyi Pohjois-Amerikassa 1800-luvulla jonkin aikaa sen jälkeen, kun omenat, vieraslaji, oli tuotu maahan. Tämä omenoita syövä populaatio syö tavallisesti vain omenoita eikä orapihlajakärpäsen historiallisesti suosimia hedelmiä. Nykyinen orapihlajaa syövä populaatio ei yleensä syö omenoita.
Joidenkin todisteiden mukaan sympatrinen lajistuminen on käynnissä. Tulikärpäset kypsyvät myöhemmin kaudella ja kypsyminen kestää kauemmin kuin omenakärpäset, ja risteytymisestä on vain vähän todisteita (tutkijat ovat dokumentoineet 4-6 prosentin risteytymisasteen).
Uuden orapihlajakärpäsen ilmaantuminen on esimerkki käynnissä olevasta evoluutiosta.
Hybridisointi
Harvoin uusi laji muodostuu, kun eri lajien yksilöt pariutuvat. Yleensä tällaisten risteytysten tuotteet ovat hedelmättömiä tai suhteellisen hedelmättömiä, joten luonnonvalinta eliminoi ne.
Yksi esimerkki uudesta ja menestyksekkäästä hybridilajista on löydetty. Uusi laji on italialaisen ja espanjalaisen varpuslinnun risteymä, ja perustiedot on tarkistettu sekvenssianalyysillä niiden veren DNA:sta. Lintu elää Italiassa alueella, jossa molemmat emolajit elävät. Se ei lisäänny espanjalaisen varpusen kanssa, vaikka se elää niiden kanssa rinnakkain.
Keinotekoinen lajisto
Uusia lajeja on luotu kotieläintalouden avulla, mutta alkuperäiset päivämäärät ja menetelmät eivät ole selvillä. Esimerkiksi kotieläiminä pidettävät lampaat on luotu risteyttämällä, eivätkä ne enää tuota elinkelpoisia jälkeläisiä Ovis orientalis -lajin kanssa, josta ne ovat peräisin.
Kotieläiminä pidettäviä nautoja voidaan sen sijaan pitää samoina lajeina kuin useita luonnonvaraisia härkä-, gaur-, jakki- ja muita lajeja, koska ne tuottavat helposti hedelmällisiä jälkeläisiä niiden kanssa.
Laboratoriolajit
Parhaiten dokumentoidut uusien lajien luominen laboratoriossa tapahtui 1980-luvun lopulla. William Rice ja G.W. Salt kasvattivat hedelmäkärpäsiä, Drosophila melanogasteria, käyttämällä sokkeloa, jossa oli kolme erilaista elinympäristövaihtoehtoa, kuten valo/pimeä ja märkä/kuiva.
Kukin sukupolvi sijoitettiin sokkeloon, ja kahdeksasta uloskäynnistä kahdesta ulos tulleet kärpäsryhmät erotettiin toisistaan, jotta ne voisivat lisääntyä omissa ryhmissään. Kolmenkymmenenviiden sukupolven jälkeen nämä kaksi ryhmää ja niiden jälkeläiset eristettiin lisääntymistarkoituksessa, koska ne suosivat elinympäristöjä voimakkaasti: ne parittelivat vain suosimillaan alueilla eivätkä parittelseet muita alueita suosivien kärpästen kanssa. Tällaisten yritysten historiaa kuvataan Rice ja Hostert (1993).
Diane Dodd pystyi myös osoittamaan, miten Drosophila pseudoobscura -lajin parittelupreferensseistä voi kehittyä lisääntymiseristystä jo kahdeksan sukupolven jälkeen, kun se käytti erilaisia ravintotyyppejä, tärkkelystä ja maltoosia.
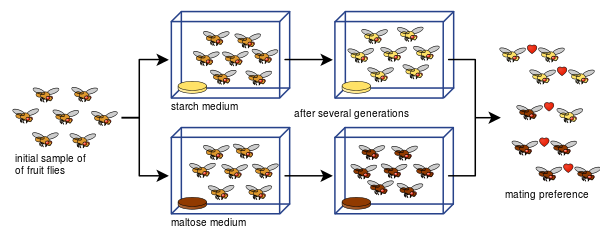
Doddin koe on ollut helppo toistaa monille muille, myös muunlaisilla hedelmäkärpäsillä ja elintarvikkeilla.
Vahvistus (Wallacen vaikutus)
Vahvistaminen on prosessi, jonka avulla luonnonvalinta lisää lisääntymiseristystä.
Sitä voi esiintyä, kun kaksi saman lajin populaatiota erotetaan toisistaan ja joutuvat sitten uudelleen kosketuksiin keskenään. Jos niiden lisääntymiseristys oli täydellinen, ne ovat jo kehittyneet kahdeksi erilliseksi yhteensopimattomaksi lajiksi.
Jos niiden lisääntymiseristys on epätäydellinen, populaatioiden väliset lisäparittelut tuottavat hybridejä, jotka voivat olla hedelmällisiä tai olla hedelmättömiä. Jos hybridit ovat hedelmättömiä tai hedelmällisiä, mutta huonompikuntoisia kuin niiden esi-isät, lisääntymiseristystä ei enää tapahdu, ja lajinmuodostus on olennaisesti tapahtunut (kuten hevosilla ja aaseilla).
Tätä perustellaan sillä, että jos hybridin jälkeläisten vanhemmilla on kummallakin luonnostaan valikoituneita piirteitä omiin erilaisiin ympäristöihinsä, hybridin jälkeläiset kantavat piirteitä molemmista, eivätkä ne sovi kumpaankaan markkinarakoon yhtä hyvin kuin kumpikaan vanhemmista. Hybridien heikko soveltuvuus aiheuttaisi sen, että valinta suosisi assortatiivista pariutumista, mikä vähentäisi hybridisaatiota.
Tätä kutsutaan joskus Wallacen vaikutukseksi evoluutiobiologi Alfred Russel Wallacen mukaan, joka ehdotti 1800-luvun lopulla, että se voisi olla tärkeä tekijä lajinmuodostuksessa.
Jos hybridin jälkeläiset ovat esi-isiään hyväkuntoisempia, populaatiot sulautuvat takaisin samaksi lajiksi sillä alueella, jolla ne ovat kosketuksissa keskenään.
Vahvistusta tarvitaan erotteluun, kun kahden lajin muodon välillä on "hybridivyöhyke". Hybridivyöhykkeet ovat alueita, joilla toisistaan poikkeavat populaatiot kohtaavat ja risteytyvät. Hybridi jälkeläiset ovat hyvin yleisiä näillä alueilla, jotka yleensä syntyvät toisistaan poikkeavien lajien joutuessa toissijaiseen kosketukseen.
Ilman vahvistamista nämä kaksi populaatiota tai lajia risteytyisivät hallitsemattomasti keskenään. Vahvistaminen voidaan saada aikaan keinotekoisissa valintakokeissa edellä kuvatulla tavalla.
Kysymyksiä ja vastauksia
K: Mitä on lajiutuminen?
V: Lajistuminen on prosessi, jossa lajit muodostuvat. Se on tärkeä osa evoluutiobiologiaa.
K: Mitä Darwin ajatteli lajien muodostumisesta?
V: Darwin ajatteli, että useimmat lajit syntyivät suoraan jo olemassa olevista lajeista, mitä hän kutsui anageneesiksi tai "fyletiseksi evoluutioksi".
K: Miten lajinmuodostukseen yleisesti suhtauduttiin 1900-luvulla?
V: 1900-luvulla tiedemiehet ajattelivat, että useimmat lajit muodostuivat aikaisempien lajien jakautuessa, mitä kutsuttiin kladogeneesiksi. Uskottiin myös, että tämä jakautuminen aiheutui tai sitä edesauttoivat eristävät mekanismit, kuten fyysinen erottuminen.
K: Miten viimeaikainen tutkimus on muuttanut käsitystämme lajinmuodostuksesta?
V: Viimeaikaiset tutkimukset ovat osoittaneet, että sukulaislajien välillä voi tapahtua risteytymistä ja että geenit voivat siirtyä näiden risteytysten kautta. Tämä tarkoittaa, että lisääntymiseristäminen ei aina ole välttämätöntä lajin määrittelemiseksi, eikä allopatriaa (fyysistä erottelua) välttämättä tarvita lajinmuodostuksen tapahtumiseen.
Kysymys: Mitä tekemistä fyysisellä erottelulla on uusien lajien muodostumisen kanssa?
V: Fyysisen erottelun kerran rinnakkain eläneiden lajien välillä uskotaan olevan tärkein tekijä uusien lajien muodostumisessa, kuten monet tieteellisessä kirjallisuudessa käsitellyt esimerkit osoittavat.
Aiheeseen liittyvät artikkelit
Tekijä
AlegsaOnline.com Lajinmuodostus: miten uudet lajit syntyvät — anageneesi, kladogeneesi Leandro Alegsa
URL: https://fi.alegsaonline.com/art/92570
Lähteet
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833